Cunabulum
Cunabulum. If you know your Latin, you might recognize this as "cradle" or "nest." If you do a Google search for it, you'll get a lot of hits for this blog, a few dictionary entries, and a helpful note that wants to know if you meant to search for
incunabulum instead. (FYI, incunabulum, according to
Wikipedia, refers to a "book, pamphlet, or broadside that was printed - not handwritten - before the year 1501 in Europe." And now you know.) Your search will also probably reveal the orchid species
Phloeophila cunabulum, so named because its flower in some ways resembles a cradle. Conspicuously missing from the search results, however, would be the application of this Latin word to an interesting morphological feature of certain species of
Stylidium, or triggerplants, a genus of small herbs mostly native to Australia.
 |
| Stylidium roseo-alatum - By Jean Hort. (Seriously, go follow her impressive photos on Flickr.) |
There are plenty of reasons to be interested in
Stylidium. For starters, the genus probably has around 300 species, which makes it the fifth largest genus in Australia. Their
variety in color and habit (ephemeral, climber, creeper) is astounding. Did I mention that they may be
carnivorous? Most notable of all, perhaps, is the irritable column or "trigger" - the fused male and female reproductive organs that hang beneath the flower and snaps into action, dusting pollinators with pollen when they come for a sip of nectar.
 |
| Stylidium debile - An animation of the column resetting after firing; each frame is 1 min. |
The complex column, a feature that many plant groups possess combined with the fact that it is one of the faster plant movements in completing its swing in
15 milliseconds is amazing enough. But if you really study this variable genus, you will find diagnostic differences in the morphology of that remarkable column. There's one particular group singled out for its interesting characteristics. I'll let
Rica Erickson, the distinguished Western Australian naturalist who passed away just a few years ago at the age of 101, describe this for us:
There is a peculiar in-folding at the apex of the Locket Triggerplant's column. It bends forward into a pouch formed by the dilatation of the column itself. It thus resembles a miniature locket with elastic hinges, enclosing the precious packet of pollen inside the lid. This can be prised apart with a pin to reveal the four neat divisions of the anthers within. ... As the stigma develops it becomes more bulky and no doubt heavy enough to cause the hinge to lose some of its elasticity and at this stage the locket hangs partly open. (Triggerplants, 1958. p. 70.) [Emphasis added]
So, the anthers develop first, then
thwack, the trigger deposits pollen on the back of visiting grey-flies. After the pollen is shed, the sticky stigma develops and pushes the anthers out of the way. The plant is now ready to receive pollen from other plants in the same manner that it just parted with its own pollen, a clever evolutionary adaptation to promote cross-fertilization.
 |
Stylidium turbinatum - Column in the set position to the left and beneath the
petals, with prominent stigma and anthers pushed to the side. The "locket" here
is somewhat open, possibly because the stigma is heavier or because of multiple firings.
Source: Holger Hennern. |
(I should mention here that a modern interpretation of species that possess a cunabulum by Australian botanist Juliet Wege excludes
S. turbinatum, noting that the column is only slightly broadened and a complete cradle for the anthers is not formed;
Wege, 2006.) With the above photo of
Stylidium turbinatum in mind, let's take a closer look at just the column of a different species, as drawn by another Australian botanist,
Allen Lowrie:
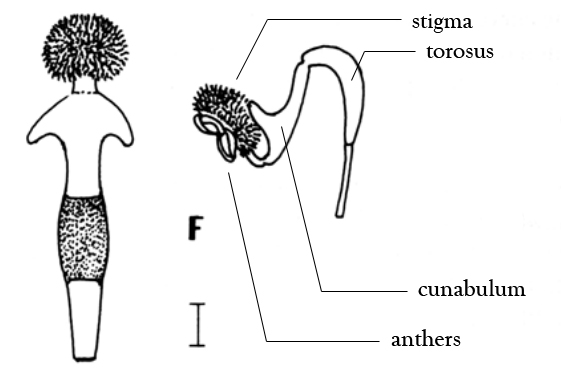
 |
Stylidium perizostera column - (Left) Looking down from
above and (right) view from the side. Scale bar = 1 mm.
Original drawing from Lowrie & Kenneally (1997), annotated by me. |
At last! The cunabulum is clearly illustrated. This particular species,
Stylidium perizostera, has lateral wings on the cunabulum. Most other cunnabula on other locket triggerplant species consist of a simple pouch. Both of the unfamiliar terms in the drawing above,
cunnabulum (Latin: cradle) and
torosus (Latin: muscular; in
Stylidium, the sensitive mobile hinge) were chosen by
Kenneally and Lowrie (1994) to be applied to the locket and hinge, respectively. Since then both words have been used only occasionally in this context, so they still remain obscure botanical terms, but they're useful when trying to use these features as diagnostic differences between species. To emphasize the special adaptation of the cunabulum, compare the backside of the widened portion of the column that corresponds to the cunnabulum in
Stylidium turbinatum and the slender, smooth column found in other species like
Stylidium purpureum.
So... what does it do?
 |
Stylidium preissii - the lizard
triggerplant, also with a cunabulum.
Photo by Jean Hort. |
You may be asking yourself, it's a cradle that holds the anthers, so what? Is there any particular function that we can infer from the morphology? Rica Erickson, again:
What is the function of this peculiar modification of the Locket Triggerplant's column? Let us watch. We must stoop very low because the plant grows close to the ground, moreover the trigger is very small. Fortunately the flies are not too wary and we can see them probing. Notice how the column suddenly shoots over. See how the force of the flying trigger flings open the locket. The anthers are held erect and pressed against the fly's shoulder. The insect flies away and swiftly the elastic hinge refolds the anthers again. The trigger has to jerk quickly to swing open the locket.
What then can be the function of the pouch? Does it conserve the moisture of the pollen grains during the dry October heat? Or is it an economical device for saving the loose grains that spill out of the anthers while waiting for insect visitors? That may be the answer, for some pouches retain a little cluster of loose grains near the hinge. (Triggerplants, 1958. p. 70.) [Emphasis added]
After Erickson's initial hypotheses of preventing desiccation or loss prevention of precious pollen, the American botanist
Sherwin Carlquist had his own impressions of the locket. One of my personal botanical heros, Carlquist
began studying Stylidium species on his trip to Western Australia in 1962 with the aid of Erickson's book which he had found in a Perth bookstore. He returned to Australia several times and identified many new species in his meticulous studies of the Australian flora. In 1969, Carlquist addressed the widened columns or "lockets" mentioned by Erickson. In his assessment, the adaptation "appears to be nothing more or less than a method of achieving self-pollination" (Carlquist, 1969). While Erickson didn't explicitly mention self-pollination, the meaning can be inferred from her suggestion that the cunabulum saves the loose pollen grains, presumably for the stigma to receive.
But as
Juliet Wege, the botanist currently working on
Stylidium, mentions, many species in this genus have
lethal post-zygotic barriers to self-fertilization. That is to say that if pollen from genetically identical individuals is received on the stigma, fertilization occurs but the embryo is aborted and viable seed is not produced. By all accounts, the floral column and trigger mechanism evolved to promote cross-pollination, further supported by the post-zygotic seed abortion found in perennial triggerplants of southern Australia. Wege concludes that, "
[i]t is therefore unlikely that the locket has evolved as a self-pollination mechanism in the perennial creeping species..." (Wege, 2006). Wege goes on to explain that tropical annual species, on the other hand, that also have a widened column may have evolved the pouch-like dilation fringed by hairs (papillae) to retain pollen and promote self-fertilization. Annuals, after all, depend on high seed set to survive from year to year.
 |
| Stylidium eriopodum - Here just to break up the wall of text. Photo by Jean Hort. |
So if the function isn't pollen retention for self-fertilization among the perennial triggerplants from southern Australia, what about the other hypotheses? Erickson's idea that it prevents desiccation of pollen during the hottest and driest months of the season has not received much mention or any data to support or refute it. Wege (2006) throws one more hypothesis into the mix by noting her field observations that pollinators, specifically long-tongued flies, will hover near recently triggered flowers to steal pollen from the anther before it has the chance to reset. Could the cunabulum provide protection from pollen pilferers? It's an attractive idea, but no published data exists.
While we're left wondering what the exact function of a dilated column, pouch, locket, or true cunabulum is, we can also ponder the evolutionary history of the trait. We have widened columns of different varieties from simple to elaborate present in multiple lineages of
Stylidium: tropical annuals, creeping perennials, tile-leaf triggerplants like
S. preissii, scale-leaf triggerplants, and whorled-leaf triggerplants. Wege is currently working on the phylogeny of the genus, but it's quite clear that this "peculiar modification" has arisen a number of times in the evolutionary history of the genus. I look forward to resolutions of such interesting questions as these.
It is amazing to me how much can be written about, and inferred from, the structure of an morphological adaptation no greater than 3 mm long on the reproductive floral column found on a genus of Australian plants. This is why botany is endlessly fascinating. This is why I love botany.
References:
Carlquist, Sherwin (1969). Studies in Stylidiaceae: new taxa, field observations, evolutionary tendencies. Aliso, 7, 13-64.
Erickson, Rica (1958). Triggerplants. Paterson Brokensha Pty. Ltd., Perth.
Kenneally, KF, & Lowrie, A (1994). Stylidium costulatum (Stylidiaceae), a new tropical species of triggerplant from the Kimberley, Western Australia and the lectotypification of S. floodii. Nuytsia, 9 (3), 343-349.
Lowrie, A, & Kenneally, KF (1997). Eight new species of triggerplant (Stylidium: Stylidiaceae) from northern Australia. Nuytsia, 11 (2), 199-217.
Wege, J (2006). Taxonomic notes on the locket trigger plants from Stylidium subgenus Tolypangium section Repentes. Nuytsia, 16 (1), 207-220.
Thanks to Paul of the Texas Triffid Ranch for the
recent mention. It provided motivation to finally get around to this overdue post. Cheers!
{kind=link}
{kind=link}
{kind=link}